Overview
Expanding the scope of protein biosynthesis
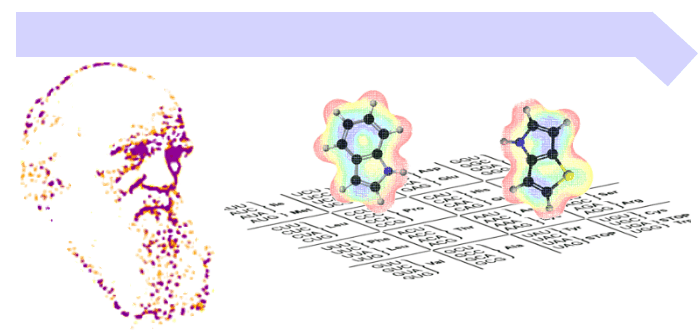
The aim of our research is to exchange, reduce or expand the number of amino acids as basic building blocks for protein synthesis, metabolism, cell structures and processes. This requires, first and foremost, the complex and radical bio-engineering with a focus on reprogramming of translation machinery by changing the coding rules, capacities and features of the genetic code.
Keep in mind following: no life forms are known to introduce amino acids into proteins beyond 20 (+2) canonical amino acids standard repertoire. However, the number of potentially novel building blocks for protein biosynthesis is virtually unlimited. This is plausible since in vitro works have demonstrated that creation of a totally new genetic code set is possible.
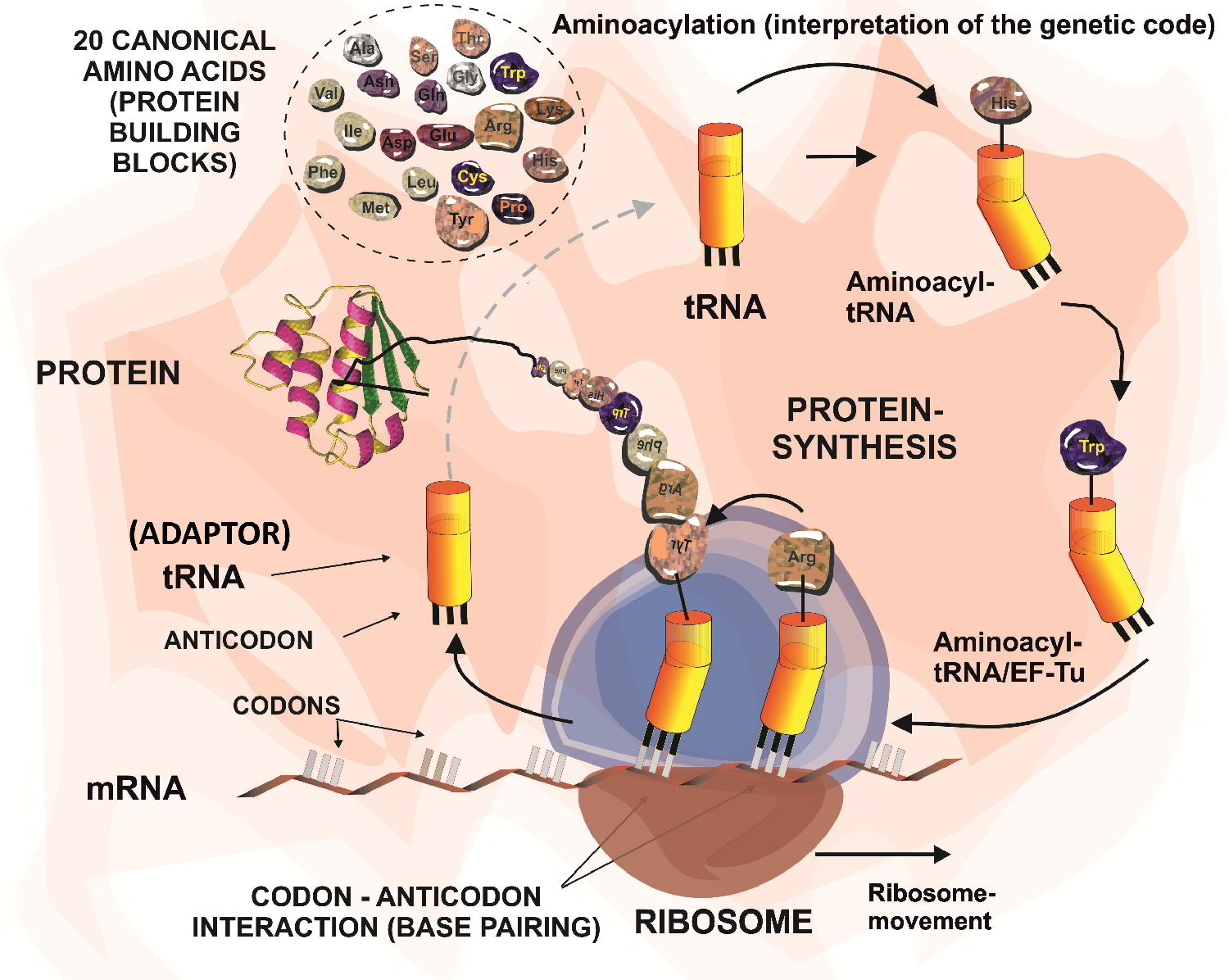
Therefore, we are very interested in studying the translational activity for the many non-canonical amino acids of natural or synthetic origin. Usually, code engineering is accomplished by use of appropriate metabolic prototypes (e.g. auxotrophies) or by direct engineering of the components of the ribosomal translation machinery (primarily aminoacyl-tRNA synthetases and tRNAs) or by a combination of both.
Genetic code engineering is aimed to explore the impact of new chemical functionalities in proteins, proteomes and whole cells. Namely, organic chemistry can provide a great diversity of non-canonical amino acids and unnatural cofactors that can be introduced into microorganisms as building blocks by experimental evolution.


Current efforts to “expand” the genetic code are in fact sophisticated in vivo protein labelling techniques primarily aimed at modifying the composition of individual proteins expressed in the frame of recombinant DNA technology. Thereby specific protein modification(s) is /are achieved by temporarily changing [re-assigning] decoding events involving only one target gene. This is the mainstream trend in this research area. However, a much more difficult task is to change (or redesign) the decoding events that are relevant to all cellular genes, resulting in a new amino acid insertion into the entire proteome.
This would require complex changes in the cell metabolic, regulatory and signaling networks in the target cells under the condition of experimentally designed long-term cultivation experiments. Clearly, this is most promising avenue towards the creation of bio-contained organism (e.g. bio-containment with genetic firewall against horizontal gene transfer). Such platforms should enable us to increase the chemical complexity, starting with a simple chemical analogues and surrogates of canonical amino acids or nucleobases, to chemically more distant and even radically different alternatives.
Despite rapid recent progress, it is not yet possible to completely alienate an organism that would use and maintain different genetic code associations permanently
Recommended reading:
Kubyshkin, V., Acevedo-Rocha, C. and Budisa N. On universal coding events in protein biogenesis. Biosystems. 2018, doi: 10.1016/j.biosystems.2017.10.004.
Kubyshkin, V. and Budisa N. Synthetic alienation of microbial organisms by using genetic code engineering: why and how? Biotechnol. J. 2017, 12 (8), doi: 10.1002/biot.201600097.
Acevedo-Rocha, C. and Budisa, N. Xenomicrobiology: a roadmap for genetic code engineering. Microb. Biotechnol. 2016, 9, 666-676, doi: 10.1111/1751-7915.12398.
Why code engineering?
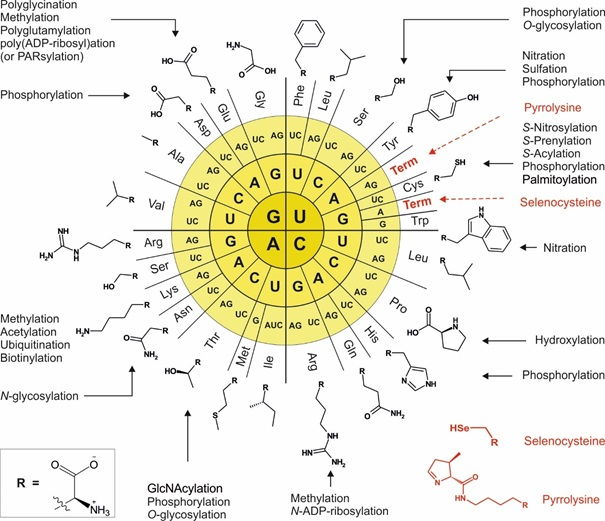
In the extant living beings, the set of 20 canonical amino acids prescribed by the universal genetic code does not span all dimensions of chemical variability necessary for diverse cellular processes advanced (e.g. metazoan) cellular forms.
Keep always in mind: only a few proteins have a final covalent structure which is a simple accurate translation of mRNA. Thus, the release of proteins from a ribosome is usually not the last chemical step in their functional maturation to become e.g. structural components of the cell or to serve as functional biochemical machines.
The majority of the proteins reach maturity and converts into active forms, either co-translationally or by post-translational processing. Indeed, evolution invented these two strategies to increase amino acid side-chain inventory. First, a small fraction of proteins is co-translationally equipped with special proteinogenic amino acids such as selenocysteine (Sec) and pyrrolysine (Pyl) by reassignment of termination codons. The second and major class of chemical modifications that contribute to the protein structure/ function diversity are post-translational modifications (PTMs). These reactions are selectively and timely coordinated events performed by dedicated enzymes or enzymatic complexes, usually in a specialized cell compartment.
Why then code engineering? Indeed, we have PTMs that encompass all the dimensions of chemical variability necessary for different cellular processes and functions.
Nonetheless, there are at least three good reasons (all good things go by three) to use code engineering.
- PTM machinery is highly complex (compartmentalization!)
- It is difficult to achieve large quantities of homogeneous product
- The recognition features for specific PTM are normally easily destroyed in laboratory settings
It is extremely difficult to mimic nature's complex machines or processes such as the PTM. Therefore, we will require genetic code engineering as long as research in microfluidic devices and systems provides us with suitably miniaturized devices or micro-devices. For example, efficiently closed membrane or semipermeable orthogonal compartments such as vesicles might act as containers (or even protocells!) with encapsulated biochemical processes.
Recommended reading: Budisa, N. - Expanded genetic code for the engineering of ribosomally synthetized and post-translationally modified peptide natural products (RiPPs). Curr. Opin. Biotechnol., 24, 591-598 (2013)
Academic application and potentials
Over the past 20 years, Ned teams have worked with various groups on joint projects. Many of these collaborations were very successful. Thereby, non-canonical amino acids proven to be valuable and useful tools for various research fields.
- Drug Design – code engineering as a route to diversify small bioactive molecules
- Biomaterials – new-to-nature congeneric peptides, proteins & complex scaffolds
- Bioorthogonal conjugations and chemistries – custom-made in vivo chemistries
- Structural Biology – noninvasive markers for 3D structures determination
- Fluorine biochemistry – fluorine as element of life and tool for NMR spectroscopy
- Spectroscopic probes in proteins – electron/proton pathways, signal transduction
- Photobiology – chromophore design (photophysics & optogenetics)
- Biophysics – protein folding, stability & energy dissipation pathways
- Biocatalysis – biocatalytic processes in cellular systems & enzymatic cascades
- Metabolic Engineering – reprograming intracellular amino acid syntheses
- Directed evolution – engineering & selection of enzymes and bacterial strains
Xenobiology, Synthetic Biology and related Technologies
Latest at the end of the 20th century it become clear that engineering of e.g. different microbial strains for fermentative production is much more efficient than classical chemical engineering. For example, Danielli predicted that “the production of sequence-determined polymers rather than the random polymers…” will create a “degree of sophistication…. far beyond thought”. In 1971 he said: “I am convinced that so many people – even biologists – aren’t aware that biology is moving from an age of analysis into an age of synthesis” [New Scientist, 1971, pp. 124].
While Danielli in 1970th was a voice in the wilderness, nowadays is well established that the main goal of Synthetic Biology (SB) is the creation of biodiversity applicable for biotechnological needs. In contrast, Xenobiology (XB) aims to expand the framework of natural chemistries with the non-natural building blocks in living cells to accomplish artificial biodiversity.
The molecules, molecular complexes and processes along the flow of genetic information ("central dogma") are particularly attractive targets for such engineering. For example, the development of alternative nucleic acids (xenonucleic acids, XNAs) based upon new base pairs, sugars and backbones. Alternatively, estranging the genetic code from its current form via systematic introduction of non-canonical amino acids should enable the development of bio-containment mechanisms in synthetic cells potentially endowing them with a ‘genetic firewall’ i.e. orthogonality which prevents genetic information transfer to natural systems.
In general, XB is an attempt to produce non-natural molecules (xenobiotic materials) using chemically modified organisms that may be endowed with a genetic firewall. In this context, processed products derived from microbes (e.g. E. coli, B. subtilis, S. cerevisiae) are particularly attractive. Such microbes are generally very suitable ‘workhorses’ for redesigning, recoding, re-inventing natural processes within short timescales.
According to Phillipe Marliere both Xenobiology and Synthetic biology have following industrial goals:
- Produce chemicals, materials and energy vectors from renewable resources.
- Design, construct and evolve microbes with novel metabolic cores and coding pathways.
- Propagate synthetic eco-systems and food-chains.
- Diversify modes of element cycling and fixation (CHNOPS + Fluorine + Silicon + Boron)
- Promote evolutionary bifurcations, prevent genetic pollution (Biocontainment, Genetic firewall).

New to-nature xenobiotic materials of future will be elaborated through combination of bio-inspired modules that will greatly surpass existing technologies with potentially high value for the society as a whole.
Synthetic Life and Biocontainment
We want to create artificial biological diversity by changing the chemical make-up of living cells to understand life, code, their origins and evolution. That’s good. We also know that the engineering of the genetic code enables the development of emergent biotechnologies. That’s fine too.But, imagine now that the mankind has set the goal to preserve current and to spread life throughout the observable universe. Once it happens, a fundamental question will be whether it would be possible to experimentally create alien life, very different than life on our planet.
This venture can greatly benefit from our knowledge of non-natural building blocks in biological systems. Xenobiology (XB) could be helpful here. We already have some ideas of how organic molecules containing boron, fluorine, silicon or other elements rarely used in earth biochemistry might be potentially beneficial to living cells that survive in artificial environments or other planetary habitat.
For example, Wowa Kubyshkin and Dirk Schulze-Makuch have speculated about the cosmic scenario in which all molecular oxygen is consumed by oxidation reactions on a planetary surface and fluorine-rich magma fluorine gas can later be released in the history of a planetary body – resulting in a fluorine-rich planetary environment. They proposed chemistries that could support life in such environments.
Even before humanity decides to take this step, we can think about the ways and consequences of our experiments, which are geared towards these specific goals. In synthetic cells with "xeno-DNA" as genetic material or alternative / different genetic codes, complete genetic isolation may work, as it aims to prevent horizontal gene transfer between species on Earth.
Consequently, the biocontainment or genetic firewall against current earth life might be an important technology with positive societal effects (as novel biosafety and biosecurity tools). Bear in mind that biocontainment implies the existence of a synthetic organism capable to survive in artificial or natural environments only by using human-supplied chemical substances (that otherwise have no biochemical source).

Future experiments will show us how far the chemical alienation of cells can be driven forward in order to create a stable barrier to information transfer between the "new synthetic" and "old biological" worlds. Ideally, a built-in "genetic firewall" could provide complete genetic isolation against horizontal gene transfer, creating the conditions for a parallel biological world with its own genetic code. This would be a “top-down” approach towards synthetic life, nicely described by De Lorenzo, Schmidt and Budisa model. They proposed a model with eight stages of development leading to the creation / emergence of synthetic life, with current research somewhere between level 5 and 6.
Recently, we proposed a sort of bottom-up approach towards synthetic life using Hartman-Smith model for the genetic code evolution in the context of RNA-World. We have rationalized the chemical logic behind the established amino acid repertoire in the extant triplet (“frozen”) code. Based on this reasoning, we were able build up a model that enabled us to predict "entry/invasion" points for noncanonical amino acids in the genetic code.

Remember, life is neither chemistry nor physics and their molecules. Life is not just information flow. Neither only energy flow. It is also not a mere self-organisation with catalytically-driven chemical supercycles. Life is more. Life is the organisation (unity) of all these phenomena. Thus, to create synthetic life with an expanded, reduced or altered genetic code, ongoing work should be combined with system bio-engineering work on self-assembled bio-orthogonal compartments and devices, along with alternative energy sources (chemo-osmotic gradients?), novel types of information transduction pathways and alternative metabolic cycles with new to nature catalytic cascades and molecular machines.
This makes a completely new biological world conceivable and plausible. The design of genetically modified organisms (in the context of classical genetics) is only the beginning of a long road in search of reliable methods for the evolution and development of artificial biodiversity while preserving the old natural world. An important task for XB, therefore, is to pursue chemically-diverse artificial evolution of viable and robust cells that can grow and replicate indefinitely in isolation from natural species.
Link for more information:
http://www.innovations-report.de/html/berichte/biowissenschaften_chemie/creating_a_parallel_biological_world_a_genetic_173974.html